摘要
早期心脏发育包括心管的形成、心管的弯曲(looping)以及心腔的形成。这些过程在所有脊椎动物中高度相似,表明心脏的基本构建蓝图在进化中得到了显著保守。从无颌类(如七鳃鳗)到人类,T-box 转录因子(如 Tbx5 和 Tbx20)在心脏形成中起关键作用,而 Tbx2 和 Tbx3 则在静脉窦-心房(sinu-atrial)和房室(atrioventricular)边界区域抑制心腔的形成。此外,不同脊椎动物的心电图特征也极为相似,尽管鱼类心脏仅有两个腔室,而哺乳动物心脏由隔膜分为四个腔室,并具有更高的心率。我们认为,哺乳动物和鸟类高性能心脏的许多特征可以追溯到变温脊椎动物心脏中较为原始的结构与功能特征。
研究要点
-
► 脊椎动物心脏的构建蓝图在进化过程中高度保守。
-
► 心脏的分子图式决定了其电生理图式。
-
► Tbx5 与 Tbx20 等转录因子对心脏形成至关重要。
-
► Tbx2 与 Tbx3 在边界区域抑制心腔形成。
-
► Tbx2 与 Tbx3 同时界定了心脏传导系统的区域。
关键词
心脏 · 传导系统 · 进化 · 发育 · 电生理 · 基因表达
1. 引言(Introduction)
在脊椎动物的进化过程中,心血管系统经历了显著的解剖学和功能性变化。最早的脊椎动物(可能以文昌鱼 Amphioxus 为代表)的心脏,可能是一个能够收缩的血管,其蠕动式运动(peristaltic movement)以低压力推动血液在血管系统中流动 [1]。随后,在鱼类中进化出了由一个有小梁(trabeculae)的心室和一个心房组成的心脏,其血液首先流经鳃部,然后流入体循环(图1)[2]。


图 1. 心脏结构与生理在进化过程中的变化
鱼类通过鳃呼吸,并且没有心脏隔膜。两栖类和爬行动物的心室未完全分隔(÷)。只有哺乳动物和主龙类(鳄鱼和鸟类)具有完整的心室隔膜(+)。与其他脊椎动物相比,哺乳动物和鸟类的新陈代谢显著提高,它们是恒温动物(红色),其心率比变温动物(蓝色)高出数倍。
体循环血压(富氧血)在恒温动物中通常高于变温动物,尽管某些高活动性的鱼类(如金枪鱼,具有类似哺乳动物的高血压。相反,肺循环血压(缺氧血)在恒温动物中较低。
缩写说明:la,左心房;lv,左心室;ra,右心房;rv,右心室;
在进化的进一步阶段,出现了具备肺循环的心脏结构,并在心脏内部实现了体循环与肺循环的部分分隔——这一特征最早出现在肺鱼(Dipnoi)中,并在两栖类尤其是爬行动物中进一步发展,其中心室出现了部分隔膜(图1)[3]。
鸟类和哺乳动物分别从类似爬行动物的祖先进化出了恒温性(endothermy),并各自独立地进化出完全隔膜的双心室结构以及厚实紧密的左心室壁,从而使体循环血压显著升高(图1)。这种系统循环与肺循环的完全分离,同时伴随着最大心率的数倍提升——这是维持恒温动物高代谢水平所需的高心输出量的关键前提 [4]。
尽管在脊椎动物心脏的进化过程中,其结构与血流动力学发生了深刻的变化,但这些变化并未在心脏整体电活动模式上表现出明显的差异。事实上,虽然恒温的鸟类和哺乳动物因高心率而使单次心动周期中的电活动总时程缩短,但所有脊椎动物的心电图都显示出一个相似的特征——即从心房最初激活到心室随后的激活之间存在一定的时间延迟(见表1,图2)。
表 1. 脊椎动物心脏的结构与遗传性传导模式。

(1) 包括七鳃鳗 [72]、软骨鱼和硬骨鱼,但不包括具有部分分隔心脏的肺鱼。
(2) 在斑马鱼中,Tbx5 在约 48 小时受精后(hpf)呈前–后梯度表达,而在其他脊椎动物中则为后–前梯度 [138]。
(3) 爬行动物和鸟类均具有 Bnf,但 Anf 仅存在于龟鳖类 [19], [51], [141]。

图 2. 心脏发育及其电活动
心管由慢速传导的原始心肌(灰色)构成,蠕动收缩产生波状心电图(ECG)。心房和心室的快速传导区(蓝色)膨出后,心电图呈现出类似成人的波形。此外,早期分腔心脏的构型与成鱼心脏相似,甚至类似于成熟的人类心脏。图中对心脏和心电图进行了示意。
然而,哺乳动物和鸟类的心脏具有结构上相似且功能上特化的心脏传导系统,该系统负责兴奋信号的产生与传播;相比之下,在爬行动物或其他任何变温脊椎动物中,并没有发现解剖学上可识别的专门心脏传导系统的证据 [5][6]。
因此,对哺乳动物和鸟类在形态和功能上极为相似的心脏却似乎独立进化这一现象的最简约解释,可能是它们共享了一个在进化上保守的心脏构建蓝图,这一蓝图为结构与功能的共同发育提供了模板。
在本文中,我们尝试回顾这一可能性,重点关注脊椎动物胚胎在早期心脏发育阶段的相似性,并强调分子层面的研究,因为分子证据在揭示同源性方面具有独特的解释力。
2. 发育中心脏的形态与心脏的进化
所有脊索动物的心脏都起源于胚胎胚盘(embryonic disk),其侧板心生中胚层(lateral cardiogenic mesoderm)在胚胎腹侧中线处汇合,形成心管(heart tube)。随后,心管发生弯曲(looping),并在外弯处膨出形成心室和心房,而心管其余部分(即内弯处)则保持管状结构(图2)[6]。
在小鼠与鸡的早期心脏发育阶段,最初阶段表现为心外前体细胞群的高增殖活性,这些前体细胞不断向心管添加细胞 [7][8]。随后,心肌细胞进入分化阶段,并伴随增殖速率显著下降;紧接着,在“膨胀阶段(ballooning phase)”中,细胞增殖再次显著活跃 [9][10][11]。有趣的是,原始的鲟鱼类(sturgeons)的发育中心脏也经历了类似的过程:最初发生一次弯曲,然后伸直,接着再经历第二次弯曲,这与小鼠和鸡的心脏发育过程相似 [12]。
虽然我们无意重新提出海克尔(Ernst Haeckel)的“个体发育重演系统发育”理论 [13],但值得注意的是,成体的原始脊索动物(如海鞘 sea squirt)在心脏形态上与早期脊椎动物胚胎的心管极为相似。此外,四足动物胚胎的心脏仅有一个带小梁(trabeculated)的心室和一个心肌外流道(myocardial outflow tract),其形态类似于鲨鱼和原始硬骨鱼类的成体心脏 [14][15][16]。进一步地,哺乳动物与鸟类胚胎的心脏在结构上与爬行动物共有一个特征:心室尚未完全分隔,而是通过心肌外流道的心室化(ventricularization)过程逐步实现部分分隔 [17][18][19]。
事实上,著名的解剖学家 Arthur Keith 曾将鲨鱼的心脏误认为是“畸形的人类心脏”[20]。在本研究中,我们采纳了进化-发育(evo-devo)学派的观点,常通过比较不同分类群在发育阶段与成体形态之间的对应关系来探讨心脏演化。
3. 心脏的电活动图式(Electrical patterning of the heart)
心肌细胞(cardiomyocytes)之间通过电连接(electrical coupling)相互作用,并具备自律性(automaticity),即它们能够产生内源性电活动节律,从而引发收缩。结果是,具有最高自发去极化速率的细胞或细胞群将决定整个心脏的节律,被称为起搏细胞(cardiac pacemaker)。
在所有脊椎动物中,优势起搏细胞均位于正在形成的心脏的流入端(inflow region) [21][22][23][24]。随着激活电流自尾侧向头侧(caudo-cranial)在心管上传播,蠕动式收缩波(peristaltic waves)推动血液向动脉极(arterial pole)**流动。
仅在发育后期几个阶段后,心室与心房腔室即可在心管上表现为局部膨出。对斑马鱼、鸡与哺乳动物的研究表明,发育中心腔的心肌表现出快速的去极化传播与几乎同步的收缩 [23][25][26][27][28]。而在静脉窦-心房边界(sinu-atrial border)、心房底部、房室管(atrioventricular canal)、心室内弯处(inner curvature)以及心肌外流道等邻近区域,心脏依然保持管状结构(图2)[6][29]。这些区域的特征是:电信号传播速度较慢、收缩持续时间较长,这一现象在鲨鱼、肺鱼和两栖类的成体心脏中也同样存在 [6][30][31][32][33][34]。
值得注意的是,在早期发育中心脏中,房室管和心肌外流道内包含心垫组织(cushions)而尚未形成瓣膜(valves),因此这些部位的缓慢而持久的收缩对于维持血液单向流动至关重要 [25][35][36][37][38]。
由于心脏中存在快、慢两类电活动区域,即使在心腔形成的早期阶段,也能记录到类似于成体心脏的心电图波形(图2)。
心脏激活动态可通过高空间分辨率的光学映射技术(optical mapping)测定,这对于研究体积极小的发育中心脏尤为有效 [39][40]。
在哺乳动物(小鼠,约E9)与鸟类(鸡,约E4)心腔形成的最早阶段,心室激活自房室管(atrioventricular canal)向心尖(apex)传播,最终延伸至流出道(outflow tract) [41][42]。
令人惊讶的是,这种激活模式与斑马鱼心脏极为相似——不仅在受精后最初几天(此时心室小梁尚未形成)[27][28],而且在斑马鱼及其他变温脊椎动物的完全发育成形的心脏中也保持这一特征(图3)。

图 3. 完全形成的心脏的心室激活模式
在斑马鱼、非洲爪蟾(Xenopus frog)和绿鬣蜥(anole lizard)(代表一般变温动物)中,心室心外膜激活(红色)始于心室基底,终止于心尖及流出道区域(蓝色)。右侧列显示所有最早(红点)和最晚激活点(白点)。
在恒温动物的成体心脏中(以人类心脏为例),激活从心腔面由基底向心尖扩展,随后扩展到心外膜面,并从心尖向基底传导。比例尺分别为:斑马鱼 0.2 mm,非洲爪蟾 1 mm,绿鬣蜥 0.5 mm。
缩写说明: lv,左心室;rv,右心室;vs,心室间隔。
在哺乳动物和鸟类中,心脏发育几天后会出现一个显著的转变:当室间隔(septum)和致密心肌层(compact wall)形成时,心室的激活首先出现在心尖部(apex)的心外膜侧(epicardial side) [43]。这种从心尖处启动的激活模式在左、右心室均源自单一激活点,因此通常被视为心室传导系统束支(bundle branches)成熟的标志 [23][42][43][44][45]。
然而,实际上这种“成熟”所代表的生物学本质尚不完全清楚,因为在此阶段,束支在解剖或分子水平上都尚未表现出明确的分化特征。也许这种激活模式的变化仅仅反映了结构性重塑(architectural changes):致密心肌壁的形成可能掩盖了早期心基部的激活信号,而室间隔的形成将早期心室的小梁网络分隔为左右两部分,分别延伸至心尖。
4. 心脏的分子图式(Molecular patterning of the heart)
除了组织结构外,心肌电信号的传播还依赖于细胞间耦合与兴奋性,这两者由离子通道蛋白的分布所决定 [46]。在心腔膨出(ballooning chambers)中,特异性表达的基因包括心房钠尿肽因子(Anf)和间隙连接蛋白 connexin40(Cx40)。间隙连接(gap junctions)是心脏功能的基础结构,使得心肌细胞之间可以通过胞质连接实现电信号的直接传播。
在哺乳动物的心室中,每个心肌细胞平均与约 11 个相邻心肌细胞通过间隙连接相连,而心房细胞平均与约 6 个细胞相连 [46][47]。
一个间隙连接由单个细胞中多个六聚体形式的连接蛋白(connexin)组成,这些蛋白与邻近细胞中的同构结构相对接。心脏中主要的连接蛋白包括 Cx30.2、Cx40、Cx43 和 Cx45(数字代表蛋白分子量大小)[48]。其中,Cx40 与 Cx43 的电导率较高,而 Cx30.2 与 Cx45 较低。Cx40 与 Cx43 精确地局限于心腔膨出区,这些区域对应于相对快速的电信号传播(图4)。

图 4. 心脏结构建构计划的分子决定因素
注意图中标注的原始环(*)。该结构仅在哺乳动物和鸡中被描述,其存在与完整心室隔膜形成同时发生。
因此,表达 Anf、Cx40 和/或 Cx43 的心肌被定义为工作心肌(working myocardium),而保持心管样表型的心肌则为原始心肌(primary myocardium)。Anf 在心房和心室中表达,而在房室管(atrioventricular canal)中不表达,这一模式在斑马鱼、非洲爪蟾、龟类及哺乳动物中均一致(表1)(鸟类及除龟类外的爬行动物中 Anf 已丢失)[19][49][50][51][52][53]。类似地,Cx40 的分布模式在蜥蜴、鸟类和哺乳动物中也保持一致 [54][48]。其他心腔膨出标志基因还包括心脏钠通道的 α 亚基 Nav1.5与钙泵 SERCA [55][56]。Nav1.5 决定动作电位上升支的速度,因此影响电信号传播速率 [46]。虽然变温脊椎动物中 Nav 的分布模式尚未完全解析,但在两栖类与爬行动物中,动作电位上升速率在传播缓慢的区室较慢,而在快速传播区室较快 [57][58]。
原始心肌(primary myocardium)的电流传播速度较慢,产生的收缩力较小,但具有较高的自发去极化率,因此能够自发收缩。这种起搏电活动(pacemaker activity)通常被认为与奇电流(funny current)有关,该电流由钾/钠超极化激活的环核苷酸门控通道(Hcn),尤其是 Hcn4 调控 [59]。然而,在完全形成的心脏中,自动节律性也可能与自发性钙释放有关 [60]。Hcn4 在整个心管中沿尾-头轴(caudo-cranial gradient)表达,但在膨出腔室中表达量下降,而在心脏流入端(即主要起搏区域)表达最强。
这一分布在鸡和人类早期心腔形成阶段尤为明显;在后期发育阶段,Hcn4 的表达逐渐局限于心脏传导系统中部区域,并在晚期胎儿阶段(late fetal stage)进一步上调 [61][62][63][64]。
在局部区域中,心房与心室腔室通过细胞增殖增强而膨出,该类心肌获得了更快的电传播速度、更低的自动性,并上调了与收缩力相关的肌节蛋白(sarcomeric proteins)表达。
因此,心脏发育的核心问题是:是什么机制决定了原始心肌与工作心肌的分区与形成?
5. T-box 转录因子决定心脏的构建蓝图
在缺乏 T-box 转录因子 Tbx20 的小鼠中,心管无法弯曲,也无法启动心腔形成;而在成年小鼠中,条件性敲除 Tbx20 的个体会在一周内死亡 [65][66][67][68]。
Tbx20 的重要性在其保守的进化表达模式中得以体现:在鸟类 [69]、爬行动物、两栖类 [70]、斑马鱼 [71] 以及原始无颌类七鳃鳗(lamprey) [72] 的心脏发育中均有表达。其他几个转录因子,如 Tbx5、Nkx2.5 和 Gata4,同样对于工作心肌表型的形成至关重要(Tbx4/5 在七鳃鳗中亦表达 [72])(图4,表1)。
值得注意的是,这些转录因子均在心脏中广泛表达,既存在于工作心肌,也存在于原始心肌 [65][66][67][73][74]。
这提示:心腔形成必须在局部受到抑制,从而维持原始心肌的存在以便在特定位置诱导腔室的形成。
在这一调控中,T-box 基因家族中两个密切相关的成员 Tbx2 与 Tbx3 发挥了关键的抑制作用 [75][76][77][78]。
6. Tbx2 与 Tbx3 抑制心腔形成
在小鼠中,Tbx2 在流入道(inflow tract)、房室管(atrioventricular canal)、流出道(outflow tract)以及心室内弯区域中表达 [75]。
除流出道外,这些区域都与小鼠心脏传导系统的发育密切相关 [48]。
重要的是,这种表达模式同样出现在人类 [79]、鸡 [80]、非洲爪蟾 [54]、斑马鱼 [81] 以及七鳃鳗 [72](表1)。
当 Tbx2 在整个心管中异常(异位)表达时,胚胎小鼠的心腔无法形成 [82]。相反,在缺失 Tbx2 的情况下,房室管心肌会启动腔室特异性基因的表达,发展出快速的电传播特征,从而破坏了正常的房室传导延迟 [83][84][85][86]。
在哺乳动物和鸟类中,Tbx3 的表达模式与 Tbx2 相似,不同之处在于:Tbx3 不在流出道中表达,并且在心室中也有表达 [54][80][87](图5)。那些被 Tbx2/3 抑制而无法分化为工作心肌的区域,其相关的基因表达显著降低,包括与肌节结构、快速电传播和钙离子调控相关的基因 [6][48][74][88]。

图 5. 胎生脊椎动物(爬行动物、鸟类和哺乳动物)中 Tbx3 的表达
图中显示了发育中心脏的腔体模型。Tbx3 表达于一个区间,从窦房边界(蓝点)延伸至整个房室管(绿色),并进入心室(虚线下方),标示出原始环,该结构在鸡和哺乳动物中后续成熟为His 束(箭头)。
在蜥蜴中,原始环是不完整的(红色虚线),因此心室隔膜未形成。绿鬣蜥的发育阶段依据 [137]。
最近的研究表明,Tbx3 能在已完全分化的心室心肌细胞中诱导出类似起搏细胞的表型 [89]。
表达 Tbx2/3 的心肌也表现出低增殖率,正如前文所述,这一特征在很大程度上可以解释心脏管环化后进一步形态塑造的机制。在小鼠中,Tbx2/3 表达的心肌在发育过程中仅缓慢增长 [48], [87],而心脏其余部分(即心房和心室)则生长更快 [11](图 6)。因此,在发育后期阶段,Tbx2/3 表达的心肌会在特定位置表现为收缩区,恰好位于窦房交界、房室管和流出道处。类似地,我们在爬行动物中也发现,房室管和流出道的心肌在早期心脏中占主要部分,但由于其生长受限,与心房和心室区相比,到了晚期阶段仅占极小比例(图 6)。
有趣的是,Tbx3 也在哺乳动物和鸟类的心室中表达,形成所谓的“原始环(primary ring)”,而这正是心室间隔形成的位置 [87](图 5)。事实上,形成双心室的关键可能并非心室间隔的产生,而是源自两个独立生长中心(即左、右心室)的扩展,这两者之间由一个抑制腔室形成的区域相隔。这与儒艮和海牛(海牛目动物)的观察结果一致——它们的左右心室几乎完全分离,只通过极少量心室肌相连,形成环状结构,类似于原始环 [90], [91](图 7)。爬行动物(如蜥蜴)则不具备原始环,也没有房室区的心室隔形成(图 5)。
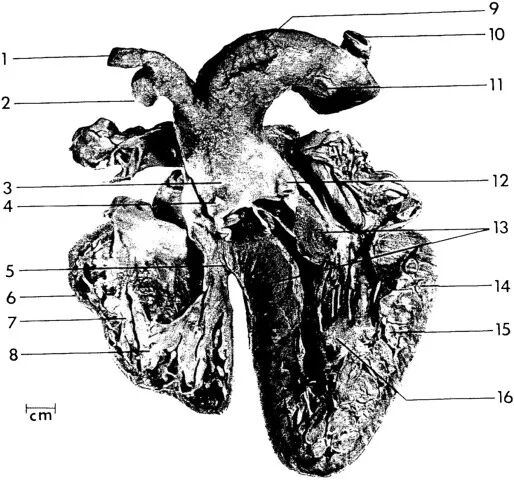
图 7. 海牛心脏的四腔视图
仅有一段非常短的心室隔(5)将左心室(13–16)与右心室(6–8)分开。这样的心室结构符合心室腔室由心管通过两个扩张点形成,中间由受抑制区域隔开的模式。
另一个 T-box 转录因子 Tbx5 在哺乳动物和鸟类的左右心室之间呈陡峭的梯度表达,对心室间隔的形成似乎具有重要作用 [19]。
目前我们对 Tbx2/3 的区域诱导与限制机制仍知之甚少,但小鼠、鸡和斑马鱼的研究表明,骨形态发生蛋白(Bmp)2 与 4 可诱导 Tbx2,甚至可能诱导 Tbx3 的表达 [80], [81], [92]。此外,Tbx2/3 可能通过诱导 Bmp2 的表达形成正反馈回路 [78](图 4)。与此一致,我们在蜥蜴中也观察到 Tbx3 与 Bmp2 的重叠表达 [54]。在更上游的调控中,斑马鱼的研究显示,Wnt 信号可驱动房室管中 Bmp4 的表达 [93],而 Foxn4 可能进一步并行诱导 Tbx2 [81]。相反,Tbx2 在房室管与心室内弯处的表达限制依赖于 Notch–Hey1/2 信号通路 [81], [94], [95] 以及 Tbx20 [96]。
因此,Tbx2/3 的重要作用在于抑制分化,使特定区域的心肌在后期能够发育为具有高自发节律与低传导速度的表型(如窦房结与房室结),或发育为具有一定自发性但快速传导特征的表型(如希氏束及其分支)。
7. 绝缘平面的发育(Development of the Insulating Plane)
在所有脊椎动物的房室沟(atrioventricular sulcus)中都可以找到纤维-脂肪组织。但只有在哺乳动物和鸟类中,这种纤维-脂肪性结构才会向胚胎房室管内延伸,形成将心室与心房绝缘的绝缘平面 [97], [98]。仅有一个区域例外,该处保留下来并发展为房室结与希氏束(AV node and His bundle) [99]–[103]。然而,这种组织的延伸常常并不完全,因此心房和心室之间可能仍通过心肌束相连 [104]。
最新研究表明,真正决定房室绝缘的关键并非绝缘平面本身的存在,而是房室心肌的分子模式化(molecular patterning) [84], [86], [105]。同样,在变温脊椎动物中,尽管在窦房区和房室区存在显著的传导延迟,但它们并不具备绝缘平面 [5], [54], [106], [107]。这强烈暗示,绝缘平面并非实现电绝缘所必需的结构,但可能提供额外的安全层。
如果绝缘平面本身并非必需,那么纤维-脂肪组织的存在是否是为了在高压差(例如心室收缩约 100 mmHg、心房约 0 mmHg)下锚定房室瓣? 金枪鱼的高压心室(约 100 mmHg)似乎否定了这一假设,因为其绝缘平面并未深入房室心肌,房室瓣直接锚定在心肌上 [107]–[109]。事实上,鳄鱼是唯一具有部分绝缘平面内侵的变温脊椎动物 [110],但即便如此,其血压、心率和血流量也远低于哺乳动物与鸟类 [2]。因此,绝缘平面的真正生理意义仍是一个未解之谜。
8. 厚壁与希-浦肯野系统的发育(Development of Compact Walls and the His–Purkinje Network)
虽然哺乳动物与鸟类的心脏源自与变温脊椎动物相同的基本“构建蓝图”,但它们的心室会形成厚实的致密壁(compact walls),这一过程始于小鼠的约 E12.5、人的约 6 周以及鸡的 HH32 阶段。致密壁停止表达 Cx40 和 Anf,但继续表达 Cx43。由于致密壁的生长速度快于腔面小梁(trabeculations),后者在心室肌中的比例逐渐缩小 [11], [48]。这与变温脊椎动物形成鲜明对比,在它们体内,小梁型心肌在成年后仍占主导地位。因此,只有在哺乳动物和鸟类中,Cx40 阳性的小梁会排列在心室腔面(图 8)。
这一由小梁型心肌向致密壁过渡的发育过程,也提供了浦肯野纤维网络形成的结构学解释。该网络定义为位于心内膜下的细薄纤维系统,仅存在于哺乳动物和鸟类。但从更广的角度看,变温脊椎动物中 Cx40/Anf 阳性的小梁心肌 可以视为浦肯野系统的同源结构。
这种分子特征也反映在电生理模式上:变温脊椎动物的小梁心室普遍自心底向心尖激活;类似地,哺乳动物和鸟类的希-浦肯野系统的激活顺序为:从心室底部的希氏束出发,经室间隔下行至心尖和浦肯野网络 [111]–[116](图 3)。随后,激活从心内膜向心外膜推进(图 3)。值得注意的是,在哺乳动物和鸟类中,仅在心外膜表面可以观察到从心尖到心底的激活方向,而这实际上是前述希-浦肯野系统自心底向心尖激活的结果。
有趣的是,小鼠心室间隔嵴上的希氏束在约 E14.5 之前仍保持 Tbx3 表达并抑制 Cx40 表达,随后在 Tbx5 的调控下,Cx40 得以逃逸抑制 [117]。
致密壁与小梁的分子差异在变温脊椎动物中尚未完全明确。在斑马鱼中,哺乳动物 Cx40 的潜在同源基因 zfCx45.6 在心室中的表达与羊膜动物相反,在致密壁中表达最强 [118]。经典的哺乳动物与鸟类心脏解剖学研究显示,心室心肌细胞的排列方向在整体上呈螺旋状(心外膜层)、环状(中层)再至螺旋状(心内膜层) [98], [119]–[121]。然而,这种结构在爬行动物及其他变温脊椎动物中并不明显,尽管其心室也可区分出不同层次 [122]–[124]。
这引发了一个问题:哺乳动物和鸟类的致密心室壁是否为独立演化的结果?我们推测,哺乳动物与鸟类心脏的致密壁可能与变温脊椎动物心脏中的结构同源(homologous)。
9. 哺乳动物与鸟类形成的传导系统差异
与变温脊椎动物相比,哺乳动物和鸟类的心脏体积更大、心率更高 [4]。除了具有希-浦系统(His–Purkinje system)之外,这两个类群还进化出了其他有助于进一步加快心室激活速度的特化结构。
例如,鸟类的心室具有一个动脉周围的浦肯野纤维网络(peri-arterial Purkinje network),该网络与冠状动脉相关联,并且同样呈 Cx40 阳性 [5], [125], [126]。更为特殊的是,鸟类甚至在心房中也存在浦肯野纤维。
相反,哺乳动物心脏的致密心室壁心肌细胞(cardiomyocytes)的宽度比其他脊椎动物大数倍——大约为 30 μm,而其他脊椎动物则不到 10 μm [127], [128]。这种心肌细胞的体积增大,尤其是宽度增加,被认为有助于加快电信号的传导速度 [46], [129], [130]。
基于心室心肌细胞的尺寸,研究者以人类的心肌生理特性为基础,对不同脊椎动物的细胞大小对传导速度的影响进行了模拟(模型系统由 90 个细胞组成,这些细胞或端对端、或侧对侧排列。膜兴奋性(以电流密度衡量)和缝隙连接的电导保持恒定;见表 2)。在其他条件相同的情况下,哺乳动物心室心肌细胞的较大宽度可使其传导速度提高 50% 以上。
这提示了一个极具启发性的可能性:
哺乳动物与鸟类可能分别通过两种不同的进化途径,实现了心室激活速度及心率的加快。
鸟类的心脏除了背侧或后部的房室结外,还具有第二个位于腹侧的房室通道——主动脉根后支(retro-aortic root branch)。该结构与希氏束(His bundle)的前部相连 [5], [125], [131]。主动脉根后支与人类先天性矫正型大动脉转位(congenitally corrected transposition)这一心脏畸形中观察到的前房室通道相似 [132]。这些特征很可能与由 Tbx3 形成的原始环的前部有关。因此,前部连接的形成可能与绝缘层的内陷(ingression)过程比与传导通路本身的发育更为密切相关。
此外,鸟类的心脏在绝缘层的心房侧还具有一个与房室结相关联的右房室环束。在哺乳动物心脏中也可以发现一个非常相似但发育程度较低的结构 [133],该结构同样表现出 Tbx3 的标记 [134]。因此,Tbx2/3 阳性的房室管和原始环被认为是正常与异常房室传导轴形成的基础。
10. 结论
尽管主要脊椎动物类群的心脏在胚胎发生的早期阶段表现出高度相似的发育过程,但它们最终发育成在结构和泵血能力上差异巨大的成体心脏。然而,在电生理层面上,心脏的激活模式却表现出高度的保守性,这种保守性反映了参与细胞间耦合的离子通道蛋白的保守分布模式。这种分布模式源自发育过程中心脏由传导缓慢的心管向快速传导的心腔转变的过程。
心腔的形成受转录因子的调控,其中 Tbx2 和 Tbx3 尤为关键,因为它们的表达可使心肌维持在心管样的原始状态,而在其缺失的区域则形成心腔(见表 1)。此外,哺乳动物和鸟类的心室间隔形成与Tbx3 阳性的间隔嵴有关,尽管 Tbx5 等其他基因也参与其中。
心室的发育始终表现为小梁状结构(trabeculated design)。在变温脊椎动物中,这种结构在成体心脏中保持不变;而在哺乳动物和鸟类中,早期的小梁状心室通过外层致密壁的扩张进一步发育。因而,在哺乳动物和鸟类中,早期的小梁最终排列于左右心室腔内壁,并分化形成希-浦系统(His–Purkinje system),从而实现心室的快速电激活。
本篇文章来源于微信公众号: 创新生命